.
En 1665 durante una de las
múltiples reuniones de la Real Academia de Ciencias de Inglaterra, Robert Hooke
describe que en un corte de corcho observado al microscopio, se observaban
pequeños espacios vacios, a los cuales les bautizo como células, la palabra
deriva del latín y significa celda, dado que estos espacios eran similares a
las celdas donde los monjes se refugian a realizar meditación contemplativa,
pues bien, Hooke es el padre de la histología y de todas las ciencias que
derivan de ella, sin embargo, Hooke supone que las células están vacías y son
instrumentos con actividad desconocida, esta conclusión no es derivada de un
sentimiento mal intencionado, sino que la tecnología en microscopia estaba aun
en pañales y las observaciones no eran tan refinadas, aunado a esto, tenemos
que las células vegetales, que componen a esa planta que llamamos corcho, están
secas y por ende no se observa más que el puro esqueleto de lo que en tiempos
pasados fue una bulliciosa célula viva.
No es hasta 1833 cuando
Robert Brown describe la presencia de una estructura circular en el centro de
las células que componen a los organismos complejos, como los mamíferos,
plantas, insectos, etc, esta estructura redonda colocada en el centro o muy
próxima del centro de la célula, se nombro núcleo y de hecho es la raíz de la
división actual de las células en forma general, sabemos que podemos tener
células eucariotas o eucariontes (Núcleo verdadero) y células procariotas o procariontes
(previo al Núcleo), al transcurrir el tiempo y acumularse las evidencias
experimentales y descriptivas se fue complementando el panorama general de las
células eucariotas, las cuales constituyen la unidad de los organismos
pluricelulares.
Una de las características
más sobresalientes de las células eucariotas, es la presencia de un sistema de
membranas interno, el cual constituye una adaptación evolutiva que a la postre
ha formado lo que en la actualidad conocemos como organelos intracelulares,
antes de pasar a describir algunos de estos organelos, me gustaría hacer un
paréntesis para mencionar que al interior de nuestra células, las sustancias se
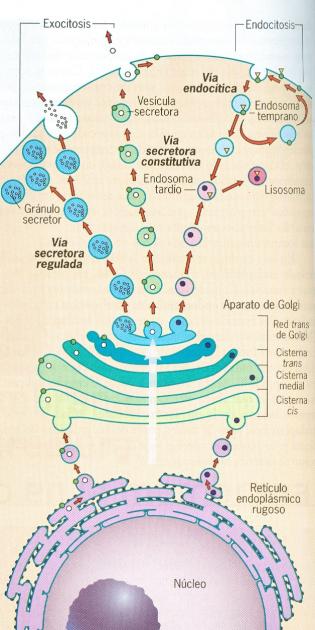
transportan mediante un sistema de vesículas.
Este sistema no es para nada
aleatorio, está regulado mediante la activación de receptores presentes tanto
en la vesícula que migra, como en el sitio de recepción en el cual será
destinado el contenido vesicular, se han descrito una gran cantidad de
proteínas que integran este sistema de reconocimiento, por cuestiones de
espacio, por esta ocasión solo mencionaremos que este complejo se conoce como
SNARE y en la vesícula que esta migrando se localiza un SNAREv o llave, el cual
interactuara con un SNAREt o cerradura, una vez que ambos complejos se
reconocen y se asocian de forma satisfactoria, la vesícula se fusiona con el
organelo receptor y libera su contenido. De esta manera se limita que ciertas
sustancias que están destinadas a secretarse en la membrana celular, terminen
en la mitocondria.
La tráfico de vesículas que
recién comentamos es conocido como vía biosintética celular, el cual lleva dos
direcciones, una anterógrada la cual va del núcleo hacia la membrana celular,
pasando por los diversos organelos, supongamos que estamos hablando de la
síntesis de una proteína, en el núcleo se sintetiza el RNA mensajero, el cual
viajara hacia el retículo endoplasmatico rugoso y mediante la acción de los
ribosomas, será traducido a una cadena polipeptídica, una serie de cadenas se
irán almacenando en una vesícula, que viajara al complejo de Golgi, donde serán
modificadas, para que adquieran una funcionalidad total, al terminar este
proceso de modificación las proteínas, serán empaquetadas de nuevo en una nueva
vesícula que las llevara hacia la membrana donde serán secretadas, para formar
el glucocáliz de la célula. La dirección retrograda corresponde al sentido
contrario, es decir, de la membrana hacia el interior. Ahora imaginemos que la
célula recibe una cantidad de prolactina, esta hormona favorece la secreción de
leche en los senos, la célula va a encapsular a la prolactina en una vesícula,
la cual viajara hacia al núcleo y activara diversos factores de transcripción
que finalizaran con la señal de producción y secreción de leche.
Conviene también hacer
mención de existen dos tipos de secreción en las células, la secreción
constitutiva que se presenta en todas las células y que es la secreción que
forma el glucocáliz y que a su vez esta recambiando los componentes de la
membrana celular, al fusionarse la vesícula que va migrando. El segundo tipo de
secreción se conoce como regulada y responde a un estímulo externo, como es el
caso de la liberación de neurotransmisores en la sinapsis o la liberación de
insulina por parte de las células beta del páncreas, que se presenta en respuesta al consumo de
alimentos ricos en carbohidratos.
En párrafos anteriores,
mencione que existen diversos organelos, los cuales tienen una función
específica, ahora comentaremos un poco sobre el retículo endoplasmatico y el
complejo o aparato de Golgi. En el caso del retículo, sabemos que es un
organelo de gran tamaño, el cual tiene continuidad con la membrana nuclear y es
básicamente una red de cisternas, del retículo podemos observar dos tipos, uno
rugoso, el cual debe este nombre a que tiene ribosomas asociados y por tal
motivo, la función principal del retículo rugoso es la síntesis de proteínas.
El retículo liso, no tiene proteínas y se ha asociado a almacenamiento de
calcio y síntesis de diversos productos, como las hormonas y ciertos lípidos.
En el caso del complejo o
aparato de Golgi, es también una red de cisternas, las cuales, tienen la
característica de ir aumentando de tamaño con dirección hacia la membrana, en
el ejemplo de la secreción constitutiva, mencione que una vesícula que migra
del retículo rugoso, llega al complejo de Golgi, a una zonda denominada como
zona de llegada o entrada, la cadena polipeptídica inicia una serie de
modificaciones postraduccionales, que la llevaran a ser funcional, esta serie
de modificaciones se va presentando al viajar entre las cisternas del complejo.
Es por esto que cada que se avanza hacia la membrana el complejo va
incrementando su tamaño, por que las proteínas van adquiriendo su configuración
estérica funcional, la cual va incrementando su tamaño, una vez que han
finalizado la serie de modificaciones, la proteína de nuevo se almacena en una
vesícula, que es liberada de la zona de salida del retículo.
Dentro de cada cisterna del
complejo de Golgi, así como, dentro del retículo, existe una gran diversidad de enzimas que realizan todas las
modificaciones, estas enzimas tienen condiciones óptimas de trabajo, pH,
concentración de iones, concentración de sustrato, etc. Estas condiciones no
serían posibles de lograr sin la existencia de una red de membranas interna,
por el momento vamos a dejar hasta aquí, en las siguientes columnas
discutiremos, lo referente a ese maravilloso mundo de la bioenergética.